More than any other factor, isolation has shaped the community
of organisms present on the Hawaiian Islands.
At least this is a reasonable prediction, if we apply principles of island
biogeography. According to this classic theory developed by ecologists Robert
MacArthur and Edward O. Wilson, species composition on islands is determined by
patterns of colonization and extinction over time. Overall species diversity on
an island is affected by its distance from a colonizing source (e.g., a
mainland) and by the island’s size. Small islands and isolated islands tend to
have lower diversity.
 |
Pacific ocean basin bathymetry/topography with the WNW to ESE trending
Hawaiian Island chain and the N to S trending Emperor Seamounts. Base
map from NOAA, NCEI. Source. |
The isolation of
Hawaii
is due to the nature of how the islands were formed geologically. The Hawaiian
chain sits in the middle of the vast Pacific crustal plate underneath the
largest ocean basin in the world. For tens of millions of years, a geologic
hotspot below the crust has continuously burped up magma to the crustal
surface, forming some of the tallest mountains in the world. Because of their
birth from an undersea hotspot underneath the Pacific plate, the islands have
never been connected to the mainland of any continent. Other hotspots dot the
planet, but
Hawaii
is remarkably distant from all other land masses. One of the
nearest islands is
Kirimati (Line Islands) at about 2000 km away; the distance from
Hilo to
San
Francisco is over 3700 km.
For terrestrial plants and animals, successful colonization
of Hawaii
came only by long-distance dispersal over thousands of kilometers of ocean. For
example, plant colonists may have had seeds that were highly tolerant of salt
water, or capable of hitchhiking on birds that landed on the islands. Seed
studies suggest that about one third of Hawaiian species arrived there by
drifting or rafting over the surface of the ocean; the remaining species
probably came with birds. For ferns (which reproduce by small spores, not
seeds) many species probably came as winds carried their lightweight spores
long distances through the atmosphere.
By looking at the kinds of native organisms present in the
terrestrial flora and fauna of a very isolated archipelago like Hawaii, we can infer
something about differences in the dispersal ability of those organisms. For
example, the native biota of the islands is missing some major groups of
animals commonly present on continents: ants, termites, reptiles and
amphibians. Mammals are also very rare in Hawaii – restricted to bats and the
endangered monk seals. These groups of organisms simply have never naturally
colonized Hawaii
because the distances are too great, the oceanic environment too hostile, or
chance was never in their factor.
 |
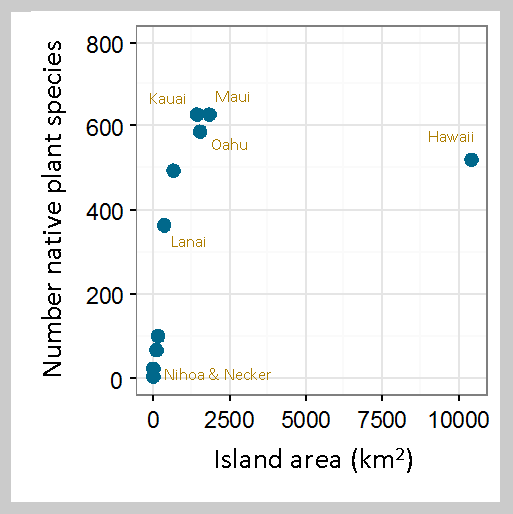
Relationship between number of native plant species and
island area for 10 of the Hawaiian Islands (8 main islands
+ Nihoa + Necker). Data are from Evenhuis and Eldredge
(2004) and Gustafson et al. (2014). The Big Island is some-
what of an outlier, but this may be reflective of its relative
youth in the island chain. |
Isolation has led to high rates of endemism on the Hawaiian Islands. Endemic species are those found in a
single location, but no where else. Today Hawaii has an estimated 1207 species of
native vascular plants, and a remarkable 88% of them are endemic to the island
chain. Some of the individual islands also have local endemics. For instance
Kauai has 219 endemic plant species while the younger islands of Oahu and Maui have 140 and 89 endemic respectively.
Isolation has affected which species have been able to reach
the islands, but other factors more local to the islands have subsequently
influenced the evolution of the successful colonists. Each island in the
Hawaiian archipelago has a life cycle of perhaps 5-20 million years from birth
to erosion and subsidence to its final days as an atoll. The
Big Island
is youngest at no more than 1 million years;
Kauai
and Nihau are the oldest of the major islands at about 6 million years old. So,
for some of the earliest colonizing lineages of organisms, their long-term
persistence on the archipelago may be due to an ability to hop from island to
island. As an older island finally sinks back into the Pacific during its old
age, species that can colonize a younger island would persist. Interestingly,
many of the species present on the islands today appear to have evolved from
colonists that arrived not more than about six million years ago (about when
Kaua’i formed) suggesting that island hopping isn’t particularly common.
 |
The honeycreeper, Vestiaria coccinea, on Acacia koa
(Fabaceae). Photo by Ludovic Hirlimann, CC BY-SA 2.0
license. Source. |
The Hawaiian biota is a good example of adaptive radiation, the
evolutionary process that results in a large and diverse group of species
diverging from a single successful colonizing species. Good examples of
adaptive radiation include the
silversword plants and the Hawaiian
honeycreepers (birds).
Studies of similarities and differences among DNA sequences
– a powerful tool to discern relationships among organisms – has also shed some
insight into the history of colonization on the islands. For the approximately 1200
native plants currently growing in
Hawaii,
it is believed that historically there were about 375 separate successful
colonizing events. Because colonization and local extinction of species on
islands is a continuous process, there were likely more successful
colonizations over the geologic history of the islands, but some of those
lineages went extinct. We can also speculate that there were probably many more
near misses where colonizing seeds or spores arrived on the shores of
Hawaii but failed to
become established, perhaps because they didn’t arrive in densities high enough
to successfully reproduce.
Of the nearly 400 plant colonizations resulting in the
contemporary Hawaiian flora, about two thirds have left us with only a single
living species. The rest have radiated into groups of related plants. For
example, the lobeliad plants now comprise 6 genera and 141 species, each
derived from what is believed to be a single colonizing event 13 million years
ago. The lobeliad radiation has resulted in a diversity of plant types from succulents
to shrubs to trees. Hawaiian radiations have sometimes occurred with little
genetic differentiation among the species but much ecological differentiation.
An example would be the ohia lehua tree (
Metrosideros),
which I wrote about in a
previous post, a species that varies considerably in
size and morphology. In contrast, other radiations can result in much genetic
diversity among a group of ecologically-similar species. For instance, a single
colonizing event for the genus
Cyrtandra
has resulted in 59 different endemic species of forest plants, all of which
live as forest understory trees or shrubs, and may therefore be pretty similar
ecologically.
Isolation and endemism – some of the features that make the
Hawaiian biota a remarkable laboratory of evolution – also are coupled with
significant threats to biodiversity on the islands. With so much unique
biological richness there is much to lose. I’ll discuss threats to Hawaiian
conservation in a later post.
References
Evenhuis NL and Eldredge LG. (eds) 2004. Natural History of
Nihoa and Necker Islands. Bishop Museum Press, Honolulu, HI,
220 pg.
Gustafson RJ, Herbst DR, Rundel PW. 2014. Hawaiian Plant
Life. Vegetation and Flora. University
of Hawaii Press, Honolulu, HI.
Price J. 2009. Hawaiian Islands,
Biology. In: Encyclopedia of Islands,
Gillespie RG and Clague DA (eds), University of California Press, Berkeley, CA,
p.397-404.














































{kind=link}
{kind=link}
{kind=link}